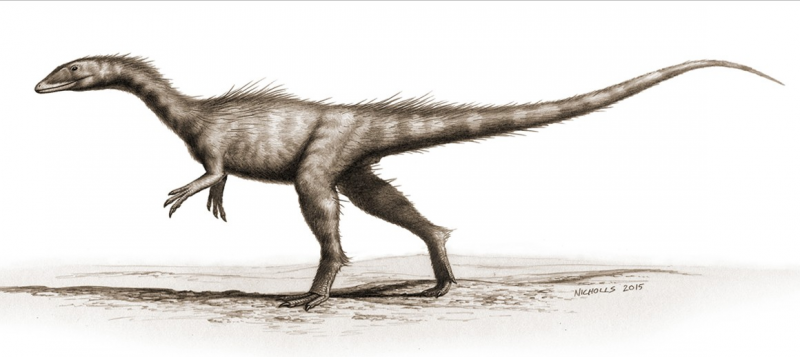
 p Dracoraptor hanigani (terópode) restaurado como um predador e necrófago da costa. Arte de Bob Nicholls (paleocreations.com). Crédito:Museu de História Natural, Londres. Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
p Dracoraptor hanigani (terópode) restaurado como um predador e necrófago da costa. Arte de Bob Nicholls (paleocreations.com). Crédito:Museu de História Natural, Londres. Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
p A evolução da endotermia (termorregulação por meios metabólicos) representa uma importante transição na história dos vertebrados. Contudo, o processo de evolução da endotermia e sua linha do tempo em pássaros e mamíferos permanecem controversos. Em um novo relatório sobre
Avanços da Ciência , Enrico L. Rezende e uma equipe de pesquisadores do Centro de Ecologia Aplicada e Sustentabilidade, e o Instituto de Ciências Ambientais e Evolucionárias do Chile, combinou um modelo de transferência de calor com dados de tamanho do corpo do terópode. Os pesquisadores então reconstruíram a evolução das taxas metabólicas ao longo da linhagem do caule da ave. Os resultados sugeriram que uma redução no tamanho constituiu o caminho de menor resistência para a evolução da endotermia, maximizando a expansão do nicho térmico, enquanto reduz os custos de requisitos elevados de energia. p Como resultado, os pesquisadores hipotetizam que o metabolismo teria aumentado com a miniaturização durante o período Jurássico Médio-Inferior (aproximadamente 180 a 170 milhões de anos atrás) para resultar em um gradiente de níveis metabólicos na filogenia dos terópodes. Embora os terópodes basais possam ter exibido taxas de metabolismo mais baixas, as linhagens não aviárias mais recentes eram provavelmente termorreguladores decentes com metabolismo melhorado. A análise forneceu uma sequência provisória no tempo das principais transições evolutivas, para o surgimento de pequenos, endotérmicos e dinossauros com penas voadores.
p A evolução da endotermia em pássaros e mamíferos é uma transição importante durante a evolução dos vertebrados, proporcionando um exemplo extraordinário de convergência evolutiva entre grupos, fundamental para sua ampla distribuição geográfica e sucesso ecológico. Embora vários grupos de invertebrados e vertebrados possam elevar suas temperaturas acima da temperatura ambiente, a capacidade de manter a temperatura corporal elevada e constante por meio da produção endógena de calor em repouso é exclusiva de pássaros e mamíferos. A observação explicou sua maior mobilidade, resistência e tolerância em uma variedade de condições. Contudo, esta estratégia é energeticamente cara e não tem nenhum vestígio virtual no registro fóssil, portanto, o ritmo e o modo de endotermia permanecem controversos durante a evolução dos vertebrados.
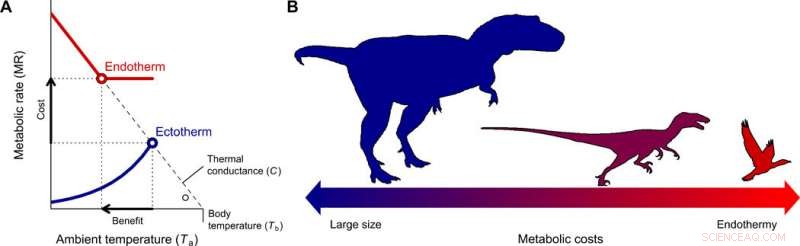 p A evolução da endotermia e miniaturização na linhagem dos terópodes levando às aves. (A) O custo-benefício para mudar de ectotermia para endotermia para diferentes faixas de tamanho corporal foi quantificado com o modelo de Scholander-Irving, que descreve como um aumento no metabolismo em repouso (custo) aumenta o nicho térmico Tb - Ta (benefício). Como não há gradiente térmico entre o organismo e o meio ambiente na ausência de produção de calor, esta curva intercepta a abcissa em Tb =Ta quando MR =0 (8). As linhas sólidas em azul e vermelho representam as curvas metabólicas de um ectotérmico e endotérmico típico, respectivamente, e os símbolos abertos representam o gradiente térmico máximo Tb - Ta possível com as taxas metabólicas de repouso, usado em nosso modelo (Eq. 2). (B) Uma redução no tamanho do corpo, consistente com aquele descrito de terópodes ancestrais a pássaros basais (22), constitui o caminho evolutivo de menor resistência, já que os custos de energia de ser grande são trocados pelos de ser endotérmico. Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
p A evolução da endotermia e miniaturização na linhagem dos terópodes levando às aves. (A) O custo-benefício para mudar de ectotermia para endotermia para diferentes faixas de tamanho corporal foi quantificado com o modelo de Scholander-Irving, que descreve como um aumento no metabolismo em repouso (custo) aumenta o nicho térmico Tb - Ta (benefício). Como não há gradiente térmico entre o organismo e o meio ambiente na ausência de produção de calor, esta curva intercepta a abcissa em Tb =Ta quando MR =0 (8). As linhas sólidas em azul e vermelho representam as curvas metabólicas de um ectotérmico e endotérmico típico, respectivamente, e os símbolos abertos representam o gradiente térmico máximo Tb - Ta possível com as taxas metabólicas de repouso, usado em nosso modelo (Eq. 2). (B) Uma redução no tamanho do corpo, consistente com aquele descrito de terópodes ancestrais a pássaros basais (22), constitui o caminho evolutivo de menor resistência, já que os custos de energia de ser grande são trocados pelos de ser endotérmico. Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
p Para entender a origem e o motivo da endotermia durante a evolução de pássaros e mamíferos, os cientistas consideraram duas questões fundamentais. Quais são os custos e benefícios desta estratégia em comparação com a ectotermia? E quais condições favoreceram uma transição para a endotermia? Rezende et al. abordou as questões usando o modelo Scholander-Irving de transferência de calor, usado para estudar termorregulação em endotérmicos por mais de 60 anos. A relação raramente era usada para ectotérmicos, mas uma vez que todos os organismos vivos produzem calor endógeno, o modelo permaneceu aplicável em estado estacionário térmico. Eles fizeram essa suposição crucial para evitar o uso de modelos dinâmicos complexos que muitas vezes são aplicados à ectotermia, o que poderia dificultar as presentes análises.
p Eles quantificaram os custos da endotermia como gasto de energia independente de massa, onde os benefícios incluíram maior mobilidade e eficiência de coleta, evasão de predadores, tolerância e colonização de uma ampla gama de condições ambientais, com aumento das taxas de crescimento e homeostase. Rezende et al. quantificou o nicho térmico que os organismos poderiam ocupar e sua expansão para estimar o benefício líquido da endotermia. Eles então calcularam o custo-benefício de adotar um estilo de vida endotérmico com referência ao ancestral ectotérmico e ao descendente endotérmico. Conforme proposto originalmente pelo biólogo evolucionista Bran K. McNab, quando os cientistas replicaram esses cálculos com estimativas exatas do tamanho do corpo, os resultados mostraram que tamanhos menores reduzem os custos de energia para evoluir para a endotermia.
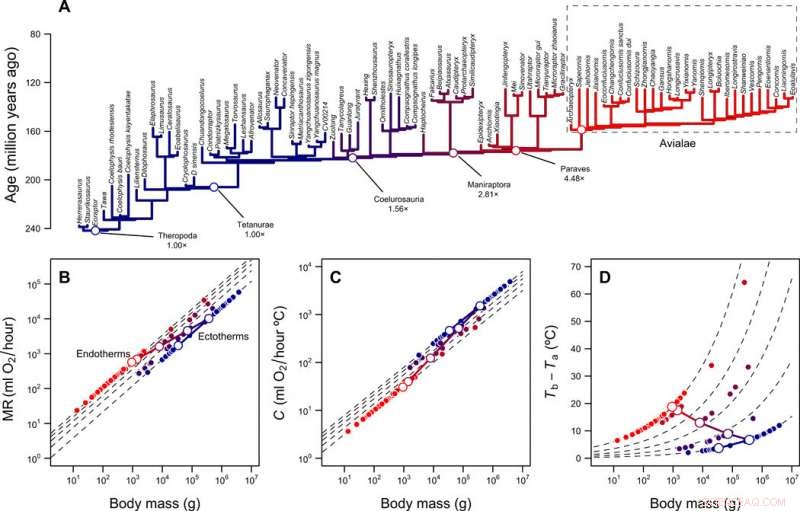 p Reconstrução dos níveis metabólicos e nicho térmico dos terópodes. (A) Filogenia de terópodes com ramos codificados por cores de acordo com os níveis metabólicos reconstruídos. (B) Escala da taxa metabólica versus massa corporal para ectotérmicos (MR =0,68 massa0,75) e endotérmicos (MR =3,4 massa0,75) e a trajetória prevista da linhagem do caule da ave durante a transição de ectotermia para endotermia. As linhas tracejadas mostram diferenças de dobra entre ectotérmicos e endotérmicos (1 × a 5 ×); símbolos abertos e fechados representam valores reconstruídos para a linhagem do caule da ave e as pontas da filogenia, respectivamente. (C) Dimensionamento da condutância térmica C e massa corporal para ectotérmicos (C =2,5 massa 0,5) e endotérmicos (C =1,0 massa 0,5), diferenças de dobra de 2,5 × a 1 ×. (D) Gradiente térmico e diferenças de dobra calculadas com a Eq. 1 (na publicação) e valores em (B) e (C). As trajetórias lineares log-log conectando MR e C do ancestral ectotérmico e do descendente endotérmico, bem como a trajetória resultante no gradiente térmico, são mostrados com as linhas contínuas. Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
p Reconstrução dos níveis metabólicos e nicho térmico dos terópodes. (A) Filogenia de terópodes com ramos codificados por cores de acordo com os níveis metabólicos reconstruídos. (B) Escala da taxa metabólica versus massa corporal para ectotérmicos (MR =0,68 massa0,75) e endotérmicos (MR =3,4 massa0,75) e a trajetória prevista da linhagem do caule da ave durante a transição de ectotermia para endotermia. As linhas tracejadas mostram diferenças de dobra entre ectotérmicos e endotérmicos (1 × a 5 ×); símbolos abertos e fechados representam valores reconstruídos para a linhagem do caule da ave e as pontas da filogenia, respectivamente. (C) Dimensionamento da condutância térmica C e massa corporal para ectotérmicos (C =2,5 massa 0,5) e endotérmicos (C =1,0 massa 0,5), diferenças de dobra de 2,5 × a 1 ×. (D) Gradiente térmico e diferenças de dobra calculadas com a Eq. 1 (na publicação) e valores em (B) e (C). As trajetórias lineares log-log conectando MR e C do ancestral ectotérmico e do descendente endotérmico, bem como a trajetória resultante no gradiente térmico, são mostrados com as linhas contínuas. Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
p A equipe explorou como este modelo de transferência de calor combinado com filogenias e reconstruções do tamanho do corpo, lançar luz sobre a evolução da endotermia em pássaros e seus ancestrais terópodes. Rezende et al. estimou os custos de evolução da endotermia ao longo da linhagem do caule da ave usando tamanhos corporais ancestrais reconstruídos, com base no registro fóssil. Para quantificar os custos de energia em cenários alternativos, eles simularam a evolução do tamanho do corpo ao longo da linhagem e obtiveram a distribuição do custo por grau no modelo. Eles assumiram um modelo não direcionado de Ornstein-Uhlenbeck (OU) entre uma massa de 10 ge 100, 000 kg com uma taxa evolutiva média equivalente àquelas relatadas para os terópodes. As simulações indicaram uma diminuição acentuada nos custos de energia por grau com a miniaturização. Os cientistas explicaram os custos reduzidos usando dois fenômenos.
p Primeiro, a expansão no nicho térmico com base em um aumento na taxa metabólica (RM) foi desproporcionalmente maior em ectotérmicos maiores devido à sua capacidade de manter uma temperatura corporal elevada (T
b ) Isso foi acompanhado por uma taxa metabólica independente de massa relativamente baixa, devido à homeotermia inicial (termorregulação estável). De acordo, eles notaram que quanto maior o tamanho inicial do ancestral ectotérmico, mais barata a transição para a endotermia. Segundo, durante a miniaturização, os animais trocaram os custos de energia de serem grandes por serem endotérmicos. Os resultados explicaram como as altas taxas de rotatividade de energia evoluíram, independentemente de seu impacto nas necessidades de alimentos e água. Apesar da variação inerente na disponibilidade de recursos no tempo evolutivo - tamanhos menores e taxas de rotatividade de energia mais altas foram sistematicamente favorecidos nesta linhagem. Resultados do trabalho concordantes qualitativamente com outros modelos na evolução da endotermia.
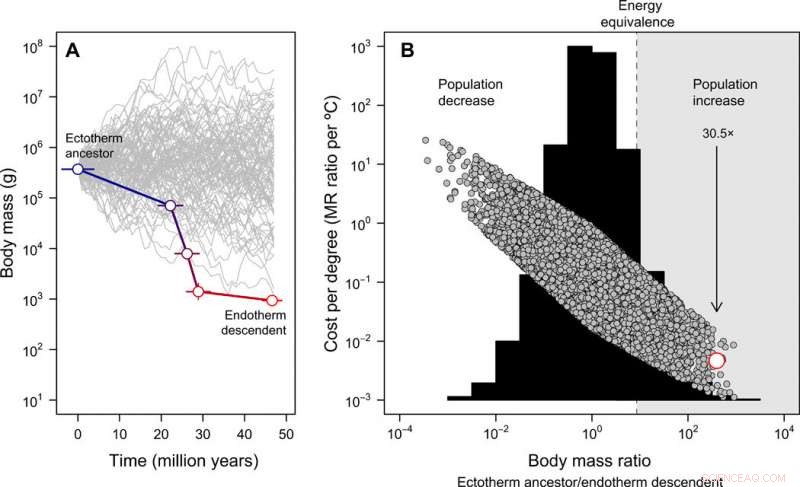 p Evolução do tamanho corporal e o custo-benefício da endotermia. (A) A miniaturização de Tetanurae para pássaros basais inferida a partir do registro fóssil, em contraste com 100 trajetórias de tamanho simuladas a partir do mesmo tamanho ancestral do corpo para fins ilustrativos (observe que, para o modelo nulo completo subsequente, o tamanho do corpo ancestral pode variar). O erro representa o SD em valores reconstruídos em 20 árvores candidatas. (B) A distribuição de frequência das razões de massa corporal obtidas em 10, 000 trajetórias simuladas do tamanho do corpo (histograma) e os custos de energia para evoluir a endotermia expressos por grau Celsius (Eq. 2 na publicação) sob este modelo nulo (símbolos cinza). Nesse caso, o tamanho ancestral do corpo foi obtido a partir de uma distribuição uniforme variando entre 10 ge 100, 000 kg. A estimativa empírica da linhagem do caule da ave é mostrada em vermelho. A região em que uma redução no tamanho do corpo compensaria os custos de energia da endotermia em evolução, possibilitando o aumento populacional em um cenário de recursos constantes, é destacado em cinza. A seta mostra o aumento esperado da dobra da população, dada a redução observada no tamanho do corpo na linhagem do caule da ave à medida que a endotermia evoluiu. Essas análises indicam que os custos de energia para evoluir a endotermia são reduzidos com a miniaturização e, como resultado, o tamanho da população pode ter aumentado apesar dos custos metabólicos de um estilo de vida endotérmico. Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
p Evolução do tamanho corporal e o custo-benefício da endotermia. (A) A miniaturização de Tetanurae para pássaros basais inferida a partir do registro fóssil, em contraste com 100 trajetórias de tamanho simuladas a partir do mesmo tamanho ancestral do corpo para fins ilustrativos (observe que, para o modelo nulo completo subsequente, o tamanho do corpo ancestral pode variar). O erro representa o SD em valores reconstruídos em 20 árvores candidatas. (B) A distribuição de frequência das razões de massa corporal obtidas em 10, 000 trajetórias simuladas do tamanho do corpo (histograma) e os custos de energia para evoluir a endotermia expressos por grau Celsius (Eq. 2 na publicação) sob este modelo nulo (símbolos cinza). Nesse caso, o tamanho ancestral do corpo foi obtido a partir de uma distribuição uniforme variando entre 10 ge 100, 000 kg. A estimativa empírica da linhagem do caule da ave é mostrada em vermelho. A região em que uma redução no tamanho do corpo compensaria os custos de energia da endotermia em evolução, possibilitando o aumento populacional em um cenário de recursos constantes, é destacado em cinza. A seta mostra o aumento esperado da dobra da população, dada a redução observada no tamanho do corpo na linhagem do caule da ave à medida que a endotermia evoluiu. Essas análises indicam que os custos de energia para evoluir a endotermia são reduzidos com a miniaturização e, como resultado, o tamanho da população pode ter aumentado apesar dos custos metabólicos de um estilo de vida endotérmico. Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
p A redução de tamanho na linhagem do caule da ave, combinou estreitamente com o caminho teórico de menor resistência para a evolução da endotermia, em seguida, Rezende et al. reconstruiu como o fenômeno pode ter se desdobrado na filogenia do terodo. O processo indicou o aumento de MR (taxa metabólica) abrangendo a maior parte do Jurássico Médio Inferior (aproximadamente 180 a 170 milhões de anos atrás), envolvendo grupos de terapeutas nos quais a ocorrência de proto-penas e penas já eram onipresentes. Eles também sugeriram taxas metabólicas altamente diversas nas linhagens concorrentes de Coelurosauria, Maniraptora e Paraves, o que poderia explicar o surgimento e diversificação desses grupos durante o período jurássico tardio. O trabalho também pode explicar a diversidade anormalmente alta de Coelurosauria em tamanhos de corpo intermediários (30 a 300 kg) em comparação com outros grupos de dinossauros. A equipe de pesquisa demonstrou ainda um modelo de preenchimento de nicho de radiação adaptativa em dinossauros do Mesozóico com taxas excepcionais de redução do tamanho do corpo na linhagem do caule da ave (ou seja, dentro de nós basais de Coelurosauria e Paraves), Contudo, eles não propuseram uma hipótese evolutiva adequada para explicar o resultado.
p Desta maneira, Enrico L. Rezende e colegas observaram dois fenômenos excepcionais durante a evolução das aves; (1) uma miniaturização sustentada (mas não necessariamente gradual) que abrange milhões de anos e (2) o surgimento da endotermia. Eles usaram reconstruções para sugerir a evolução concomitante da endotermia com uma diminuição no tamanho ao longo da linhagem do caule da ave, como também proposto anteriormente para mamíferos. Os clados de terapeutas correspondentes exibiram todo um espectro de MR. As interpretações indicam que a endotermia precedeu a evolução do vôo e a redução acentuada no tamanho do corpo durante a evolução da linhagem do caule das aves foi acompanhada por uma grande mudança nos níveis metabólicos.
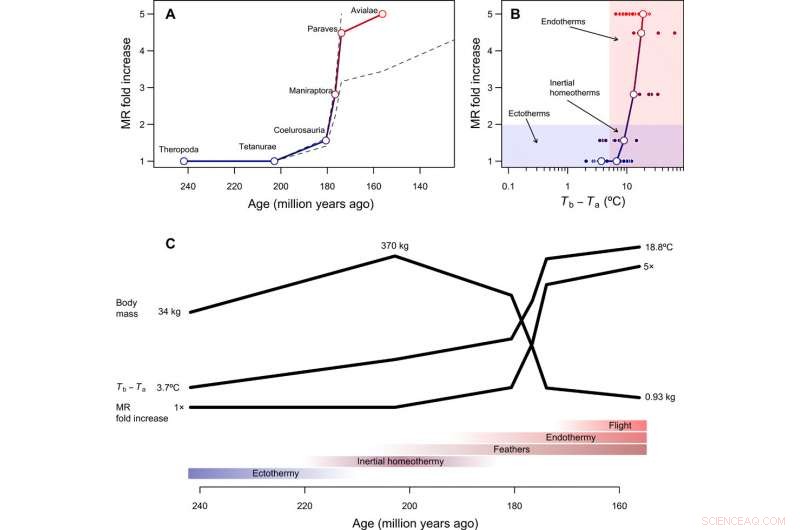 p Tempo e modo na evolução da endotermia. (A) Curso temporal reconstruído da evolução metabólica na linhagem do caule da ave, com linhas tracejadas mostrando como as reconstruções mudam assumindo que os Paraves ou Neornithes eram totalmente endotérmicos em vez do pássaro basal [para cálculos com Neornithes, assumimos um tamanho corporal de 150 g com base em estimativas para Vegavis e uma estimativa de tempo de 100 Ma atrás]. O aumento na RM foi calculado dividindo-se a RM reconstruída durante a transição para a endotermia pela RM esperada para um ectotérmico de tamanho semelhante e é, portanto, adimensional e independente do tamanho do corpo. (B) O caminho evolutivo de menor resistência da ectotermia à endotermia inclui a homeotermia inercial como um estágio de transição, seguido por um aumento no metabolismo concomitantemente com uma redução no tamanho. (C) Sequência hipotética de transições evolutivas na linhagem do caule da ave, que combina os resultados deste estudo com reconstruções filogenéticas de estruturas epidérmicas e capacidade de voo ativo (ver o texto principal). Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
p Tempo e modo na evolução da endotermia. (A) Curso temporal reconstruído da evolução metabólica na linhagem do caule da ave, com linhas tracejadas mostrando como as reconstruções mudam assumindo que os Paraves ou Neornithes eram totalmente endotérmicos em vez do pássaro basal [para cálculos com Neornithes, assumimos um tamanho corporal de 150 g com base em estimativas para Vegavis e uma estimativa de tempo de 100 Ma atrás]. O aumento na RM foi calculado dividindo-se a RM reconstruída durante a transição para a endotermia pela RM esperada para um ectotérmico de tamanho semelhante e é, portanto, adimensional e independente do tamanho do corpo. (B) O caminho evolutivo de menor resistência da ectotermia à endotermia inclui a homeotermia inercial como um estágio de transição, seguido por um aumento no metabolismo concomitantemente com uma redução no tamanho. (C) Sequência hipotética de transições evolutivas na linhagem do caule da ave, que combina os resultados deste estudo com reconstruções filogenéticas de estruturas epidérmicas e capacidade de voo ativo (ver o texto principal). Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
p O cenário proposto explica por que os terópodes sistematicamente favorecem um estilo de vida caro (maior taxa de rotatividade de energia), apesar de seus custos de energia, e miniaturização sustentada precedendo a origem dos pássaros. Além disso, um estado de homeotermia inercial constituiu um estado de transição necessário com baixos custos metabólicos. Se grandes terópodes ancestrais estivessem fisiologicamente comprometidos com a homeotermia, Rezende et al. então espere que eles tenham se afastado da alometria metabólica ectotérmica (relação do tamanho do corpo com a forma, anatomia, fisiologia e comportamento) com miniaturização. Durante o tempo evolutivo, é razoável esperar que as linhagens explorem nichos recém-abertos e, eventualmente, se diversifiquem. Os resultados deste trabalho são preliminares; eles simplesmente destacam que é provável que haja mais na evolução da endotermia em arcossauros, dinossauros e pássaros atuais. Os resultados formam uma hipótese de trabalho que pode em estudos futuros ser combinados com análises anteriores sobre a evolução do tamanho e outras características na linhagem do caule das aves para formar interpretações bem definidas de sequências temporais durante as principais transições evolutivas. p © 2020 Science X Network