Crédito:Wikipedia commons
Um dos maiores desafios da biologia hoje é explicar a estrutura das cristas, as membranas internas das mitocôndrias. Uma explicação neste caso é um conjunto de princípios para prever que forma as cristas terão após manipulações metabólicas básicas do ambiente em que as mitocôndrias estão. mal imaginado.
Avanços recentes em técnicas como microscopia de luz de super-resolução de células vivas e tomografia eletrônica deram uma nova visão sobre o comportamento dinâmico das cristas. Uma arquitetura detalhada de todo o volume mitocondrial agora pode ser construída a partir de uma série de imagens de inclinação que são retroprojetadas para criar tomogramas 3D. Na segunda-feira, discutimos como as cristas são transformadas e reconfiguradas de acordo com a abundância e a saúde de várias proteínas da membrana interna e da matriz. O pretexto para essa análise foram as semelhanças estruturais e bioquímicas entre as membranas das mitocôndrias, tilacóides e mielina que, presumivelmente, ajudam a canalizar metabólitos na produção de energia.
Em um artigo recente do
Open Biology da Royal Society , os pesquisadores explicam a biogênese das cristas através das atividades coordenadas de quatro vias principais que são evolutivamente preservadas, desde protistas e leveduras até eucariotos superiores como nós:formação de dímeros e oligomerização da ATP sintase nas bordas das cristas, montagem do 'sítio de contato mitocondrial e sistema organizador de cristas' (MICOS) nas junções de crista, remodelação da membrana por uma GTPase associada à dinamina associada à membrana interna (Mgm1 em levedura e OPA1 em mamíferos) e ajuste adequado da composição lipídica da membrana.
Para a primeira via envolvendo a ATP-sintase, várias coisas são aparentes. Como relatamos anteriormente, a dimerização espontânea da ATP-sintase em ângulos precisamente definidos e dependentes da espécie em fileiras ordenadas dita a geometria do piso térreo. Em contraste com os complexos respiratórios I-IV, que são montados na membrana limite interna plana, a ATP-sintase (complexo V) é totalmente montada profundamente nas membranas das cristas. Enquanto várias proteínas da subunidade ATP-sintase são dispensáveis para a formação adequada de cristas, as subunidades Atp20 e Atp21 são estritamente necessárias.
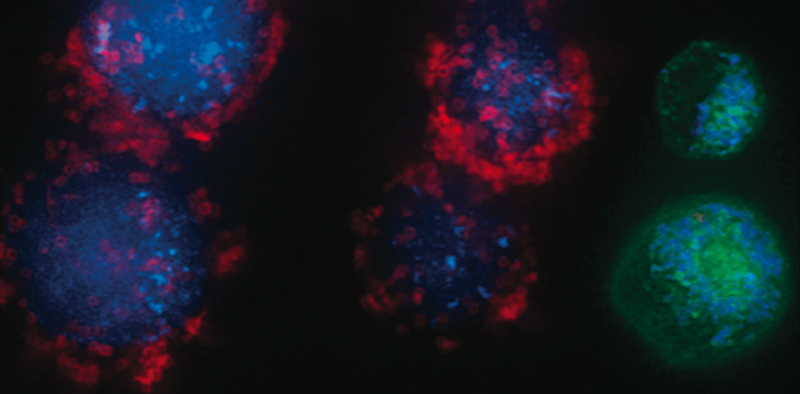
Um excesso de ADP induz uma conformação condensada com grandes espaços intracristais inchados. Em contraste, sob condições limitantes de ADP, as mitocôndrias adotam a conformação ortodoxa com espaço intracristal contraído. Na ameba gigante Chaos carolinensis, as mitocôndrias normalmente contêm cristas tubulares orientadas aleatoriamente. Com a fome, as cristas aumentadas adotam uma morfologia cúbica com um padrão em ziguezague. Em camundongos, agentes apoptóticos causam a fusão de cristas individuais com subsequente liberação de citocromo c do espaço intracristal para a região limítrofe.
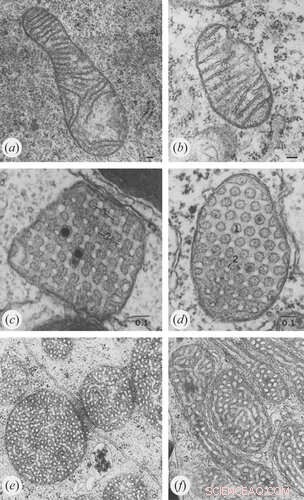
Crédito:Klecker e Westermann, Biologia Aberta (2021). DOI:10.1098/rsob.210238
Para a segunda via, a montagem de sítios de contato MICOS, a pesquisa determinou que proteínas críticas, como as da família de genes relacionados ao MIC60, estão presentes desde os ancestrais endossimbióticos das mitocôndrias – as α-proteobactérias. Muitos desses ancestrais mitocondriais já apresentam estruturas de membrana intracitoplasmática diferenciadas. Invariavelmente, as espécies que simplificaram suas mitocôndrias ao ponto em que as cristas estão ausentes correspondentemente não possuem os genes relacionados ao MICOS. A re-expressão de homólogos de MIC60 em mutantes de levedura Δmic60 resgata os defeitos mitocondriais ultraestruturais.
A terceira via inclui as GTPases relacionadas à dinamina, que coordenam a fusão e a fissão das membranas interna e externa. Na fissão, essas proteínas se polimeizam em anéis contráteis que aplicam forças constritivas para comprimir as mitocôndrias. O resultado é agora entendido como dependente das interações dessas proteínas, tanto com o complexo MICOS e junções de cristas, quanto com os sistemas de transporte de membrana interna e externa que se reúnem lá. Estes incluem os complexos translocadores de membrana TIM e TOM.
A quarta via compreende os próprios fosfolipídios da membrana mitocondrial. As mitocôndrias abrigam a via biossintética da cardiolipina e também estão envolvidas na síntese de fosfatidiletanolamina. Juntamente com a fosfatidilcolina, estes são os três principais fosfolipídios com os quais as mitocôndrias estão trabalhando. A maioria dos blocos de construção dos lipídios mitocondriais é sintetizada no RE e, portanto, deve ser importada por mecanismos que envolvem aposição próxima ao RE. Uma vez dentro da membrana externa, a distribuição de lipídios é mediada por proteínas de transporte localizadas no espaço intermembranar da família Ups/PRELI,
As mitocôndrias não criam geometria a partir do zero, mas sim aproveitam e constroem as formas físicas naturais que ocorrem espontaneamente nos lipídios. Deixados por conta própria, os lipídios formam estruturas lamelares concêntricas que podem ser expandidas e aumentadas por proteínas específicas. Medições em escala fina revelaram agora que as cristas individuais são funcionalmente independentes e podem ter potenciais de membrana significativamente diferentes.
A formação de Cristae envolve uma interação estreitamente conectada das quatro influências de modelagem acima. Por exemplo, as atividades do complexo MICOS e da dimerização da ATP sintase são cooperativas e antagônicas. MICOS induz curvatura negativa da membrana enquanto a ATP sintase induz curvatura positiva nas pontas e bordas das cristas. New computational models, as currently under development in labs throughout the world, where ratios of these different components can be finely tweaked and adjusted, will greatly aid in defining what controls the form of mitochondria.