Crédito:Wikipédia
O cobalto fica no centro do anel de corrina da vitamina B12 e das importantes cobalaminas que derivamos dele. Talvez surpreendentemente, apenas duas de nossas enzimas se preocupam em usar esses cofatores dolorosamente construídos e meticulosamente canalizados. Por que nossas células se esforçam tanto para obter um pouco da magia do cobalto, e quais propriedades catalíticas podem torná-la tão especial?
Outros metais essenciais incomuns, como molibdênio, selênio e iodo, são usados de forma semelhante apenas com moderação nas células, e ainda assim mantemos a capacidade de sintetizar completamente todos os derivados úteis para esses elementos. Para domar o molibdênio, construímos um cofator de molibdopterina elaborado, enquanto para aproveitar o iodo, montamos a tiroxina. Para incorporar o selênio nas poucas selenoproteínas que o requerem, a elaborada maquinaria SECIS embaralha o código de mRNA para atrair um tRNA único, sobre o qual sua carga de cisteína é transformada em selenocisteína. Em cada um desses casos, os pesquisadores entendem as propriedades especiais dos metais envolvidos que os tornam indispensáveis.
Por exemplo, comparado ao enxofre, o selênio é um nucleófilo melhor que reagirá com espécies reativas de oxigênio mais rapidamente, mas sua falta de caráter de ligação π significa que ele também pode ser reduzido mais facilmente. Selenoproteínas como GPX4 (glutationa peroxidase) são correspondentemente mais resistentes tanto à superoxidação quanto à inativação irreversível. Da mesma forma, a exigência inelutável de molibdênio, um composto redox de dois elétrons que pode alternar entre os pares redox +4/+5 e +5/+6, reflete várias habilidades não tão comuns. Ele pode realizar reações redox diversas e energeticamente desafiadoras; pode atuar como um sumidouro ou fonte de elétrons em baixo potencial redox; e (junto com o tungstênio muito mais raro) pode efetivamente transferir átomos de oxigênio e enxofre durante reações que ocorrem em baixo potencial.
Uma tentativa notável de adivinhar o caráter essencial do cobalto foi avançada em um comentário recente no
PNAS pelo extraordinário geoquímico Michael Russell. Posicionado entre Fe e Ni na tabela periódica, Russell observa que "o elemento é particularmente 'energicamente denso' com elétrons emparelhados na órbita externa. Sua ocorrência como uma liga metálica em serpentinitos com uma valência variável que se estende de Co
+
até Co
4
+
, seus vários estados de spin e suas conformações contrastantes o tornam único, com incontáveis contribuições a serem feitas à eletrônica, à catálise e ao surgimento da vida. De fato, a cooperação Co-Fe acaba de ser investigada na extremidade oposta do espectro redox - a eletrocatálise do O
2 reação de evolução. As substituições de Co são inviáveis, como no metabolismo e em algumas catálises de átomos duplos, ou estão em um futuro um tanto remoto."
Os comentários de Russell são em resposta a um artigo anterior de He et. al. que demonstraram que a redução hidrotérmica de bicarbonato em hidrocarbonetos de cadeia longa (≤24 carbonos) é possível através do uso de metais ferro e cobalto. Essas descobertas potencialmente explicam tanto a origem abiogênica do petróleo quanto os principais eventos no surgimento da vida. Uma vez que restos de porfirinas e corrinas que são críticas para a vida podem ser encontrados em meio a depósitos de petróleo, uma questão crítica se torna:a vida inventou essas moléculas, ou eles primeiro usaram fac-símiles abióticos dessas moléculas e só mais tarde desenvolveram a capacidade concomitante de sintetizá-las? para eles mesmos?
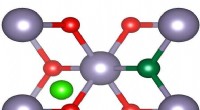
A resposta à minha pergunta de Russell foi que, em sua opinião, a vida provavelmente inventou a coordenação semelhante à corrina por meio de um motivo de quatro aminoácidos-peptídeo glicina-glicina-histidina capaz de aprisionar o átomo de cobalto. Curiosamente, as porfirinas, que abrigam ferro ou cobre em seus centros, e clorinas, que fazem o mesmo com o magnésio, devem ser contraídas em corrinas para ligar o cobalto. Essa especificidade aparentemente ocorre apesar dos raios atômicos quase idênticos (por volta de 125 pm) para a formação elementar contígua Fe, Co, Ni, Cu. Na visão de Russell, o cobalto (e outros metais de transição) necessários no surgimento da vida estavam ativos em depósitos do mineral ferrugem verde, também conhecido como fougerita, em fontes hidrotermais alcalinas. O corrinóide de cobalto unido a aglomerados de ferro-enxofre formam o coração das vias primitivas de acetil coenzima-A dos acetogênios e dos metanógenos que se encontram na base de nossa árvore evolutiva. Esta proteína Co(FeS) medeia a ligação ou separação de um grupo metil para ou de monóxido de carbono ou outra entidade envolvida na biossíntese de acetil-CoA.
A forma de vitamina B12 usada pela nossa enzima metilmalonil-CoA mutase localizada nas mitocôndrias para a quebra de ácidos graxos e aminoácidos é conhecida como adenosilcobalamina (AdoCbl). A outra enzima que utiliza cobalamina, a metionina sintase, atua no citosol e usa um cofator de metilcobalamina em que o grupo adenosil é substituído por um grupo metil. Plantas terrestres e fungos não sintetizam ou requerem cobalamina, pois não possuem metilmalonil-CoA mutase e possuem diferentes tipos de metionina sintase que não requerem B12. Quando essas enzimas não estão funcionando adequadamente, suas moléculas precursoras podem, presumivelmente, atingir níveis elevados, causando problemas como doenças desmielinizantes e anemia perniciosa.
Embora a estabilidade térmica e a alta densidade de energia do cobalto o tornem um componente ideal para os cátodos das baterias de lítio, sua utilidade para a vida vem de suas muitas outras propriedades únicas, algumas descobertas e outras ainda a serem encontradas.