De acordo com as leis básicas da física, todos os seres vivos precisam de alguma forma de energia do ambiente para sustentar a vida. Claramente, diferentes organismos desenvolveram vários meios de coletar combustível de várias fontes para alimentar o maquinário celular que conduz processos cotidianos como crescimento, reparo e reprodução.
Plantas e animais obviamente não adquirem alimentos (ou seu equivalente em organismos que não podem realmente "comer" nada) por meios semelhantes e suas respectivas entranhas não digerem remotamente as moléculas extraídas de fontes de combustível da mesma maneira. Alguns organismos precisam de oxigênio para sobreviver, outros são mortos por ele, e outros ainda podem tolerá-lo, mas funcionam bem em sua ausência.
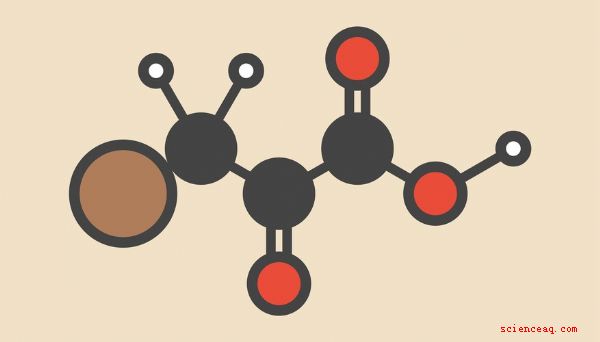
Apesar da variedade de estratégias que os seres vivos empregam para extrair energia das ligações químicas do carbono compostos ricos, a série de dez reações metabólicas denominadas coletivamente glicólise Uma visão geral dos principais insumos e produtos da glicólise é um bom ponto de partida para entender como as células convertem as moléculas reunidas do mundo externo em energia para sustentar os inúmeros processos vitais nas quais as células do seu corpo estão continuamente envolvidas. Os reagentes da glicólise são frequentemente listados glicose e oxigênio, enquanto a água, o dióxido de carbono e o ATP (adenosina trifosfato, a molécula que mais costuma usar para alimentar processos celulares) são fornecidos como produtos de glicólise, da seguinte forma: Chamar isso de "glicólise", como alguns textos, está incorreto. Esta é a reação final da respiração aeróbica como um todo, da qual a glicólise é o passo inicial. Como você verá em detalhes, os produtos da glicólise em si são, na verdade, piruvato e uma quantidade modesta de energia na forma de ATP: O próximo passo é a conversão de glicose-6-fosfato em frutose-6-fosfato (F-6- P) Isso é simplesmente um rearranjo de átomos, ou uma isomerização, sem acréscimos ou subtrações, de modo que um dos átomos de carbono dentro do anel de glicose seja movido para fora do anel, deixando um anel de cinco átomos em seu lugar. (Você deve se lembrar que a frutose é "açúcar da fruta", um elemento alimentar comum e natural.) A enzima que catalisa essa reação é a fosfoglucose isomerase. O terceiro passo é outra fosforilação, catalisada pela fosfofructoquinase (PFK) e produzindo 1,6-bifosfato de frutose (F-1,6-BP). Aqui, o segundo grupo fosfato é unido ao átomo de carbono que foi retirado do anel na etapa anterior. (Dica da nomenclatura química: A razão pela qual essa molécula é chamada de "bifosfato" em vez de "difosfato" é que os dois fosfatos são unidos a diferentes átomos de carbono, em vez de um ser unido ao outro oposto à ligação carbono-fosfato. bem como a etapa de fosforilação anterior, o fosfato fornecido vem de uma molécula de ATP, portanto essas etapas iniciais da glicólise requerem um investimento de duas ATP. O quarto passo da glicólise quebra uma molécula de seis carbonos agora altamente instável em duas moléculas de três carbonos diferentes: 3-fosfato de gliceraldeído (GAP) e fosfato de di-hidroxiacetona (DHAP). A aldolase é a enzima responsável por essa clivagem. Você pode discernir pelos nomes dessas moléculas de três carbonos que cada uma delas obtém um dos fosfatos da molécula mãe. Com a glicose sendo manipulada e dividida em aproximadamente igual peças devido a uma pequena entrada de energia, as reações restantes da glicólise envolvem a recuperação dos fosfatos de uma maneira que resulta em um ganho líquido de energia. A razão básica para que isso ocorra é que a remoção de grupos fosfato desses compostos é mais energética do que simplesmente levá-los diretamente das moléculas de ATP e aplicá-los a outros propósitos; pense nos passos iniciais da glicólise em termos de um velho ditado - "Você também precisa gastar dinheiro para ganhar dinheiro." Como G-6-P e F-6-P, GAP e DHAP são isômeros: Eles têm a mesma fórmula molecular, mas estruturas físicas diferentes. Por acaso, o GAP está na via química direta entre glicose e piruvato, enquanto o DHAP não. Portanto, na quinta etapa da glicólise, uma enzima chamada triose fosfato isomerase (TIM) se encarrega e converte DHAP em GAP. Essa enzima é descrita como uma das mais eficientes em todo o metabolismo energético humano, acelerando a reação que catalisa em um fator de aproximadamente dez bilhões (10 <10). Em etapa oito, o 3-PG é convertido em 2-fosfoglicerato (2-PG), graças à fosfoglicerato mutase, que extrai o restante grupo fosfato e o move um carbono. As enzimas mutase diferem das isomerases, pois, em vez de reorganizarem significativamente a estrutura de uma molécula inteira, elas apenas transferem um "resíduo" (neste caso, um grupo fosfato) para um novo local, deixando intacta a estrutura geral. No passo nove, no entanto, essa preservação da estrutura é discutida, pois o 2-PG é convertido em piruvato de fosfoenol (PEP) pela enzima enolase. Um enol é uma combinação de um alceno e um álcool. Alcenos são hidrocarbonetos que incluem uma ligação dupla carbono-carbono, enquanto álcoois são hidrocarbonetos com um grupo hidroxila (-OH) anexado. O -OH no caso de um enol está ligado a um dos carbonos envolvidos na ligação dupla carbono-carbono do PEP. Finalmente, na décima e última etapa da glicólise, o PEP é convertido em piruvato por a enzima piruvato quinase. Se você suspeitar dos nomes dos vários atores nesta etapa que outras duas moléculas de ATP são geradas no processo (uma por reação real), você está correto. O grupo fosfato é removido do PEP e anexado ao ADP à espreita nas proximidades, produzindo ATP e piruvato. O piruvato é uma cetona, ou seja, possui um carbono não terminal (isto é, um que não está no final da molécula) envolvido em uma ligação dupla com oxigênio e duas ligações simples com outros átomos de carbono. A fórmula química do piruvato é C 3H 3O 4O 3, mas a expressão como (CH3) CO (COOH) oferece uma imagem mais esclarecedora do produto final da glicólise. A quantidade total de energia liberada (é tentador, mas errado dizer "produzido", como "produção de energia" é um termo impróprio) é convenientemente expressa como dois ATP por molécula de glicose. Mas, para ser mais preciso matematicamente, isso também é 88 quilojoules por mole (kJ /mol) de glicose, igual a cerca de 21 quilocalorias por mole (kcal /mol). Uma toupeira de uma substância é a massa dessa substância que contém o número de moléculas de Avogadro, ou 6,02 × 10 23 moléculas. A massa molecular da glicose é de pouco mais de 180 gramas. Como, como observado anteriormente, a respiração aeróbica pode derivar bem mais de 30 moléculas de ATP por glicose investida, é tentador considerar trivial a produção de energia da glicólise como trivial. , quase inútil. Isso é completamente falso. Considere que as bactérias, que existem há quase três bilhões e meio de anos, podem sobreviver muito bem usando apenas a glicólise, porque essas são formas de vida requintadamente simples que têm poucos requisitos que os organismos eucarióticos fazem. De fato, é possível visualizar a respiração aeróbica de maneira diferente, mantendo todo o esquema de cabeça para baixo: embora esse tipo de produção de energia seja certamente uma maravilha bioquímica e evolutiva, os organismos que a utilizam em sua maior parte absolutamente dependem dela. Isso significa que, quando não há lugar para encontrar oxigênio, organismos que dependem exclusiva ou pesadamente do metabolismo aeróbico - ou seja, todo organismo que lê esta discussão - não podem sobreviver por muito tempo na ausência de oxigênio.
é comum a praticamente todas as células, tanto em organismos procarióticos (quase todos os quais são bactérias) quanto em organismos eucarióticos (principalmente plantas, animais e fungos).
Glicólise: reagentes e produtos
C 6H 6O 12O 6 + 6 O 6 + 6 O 2 -> 6 CO 2 + 6 H 2O + 36 (ou 38) ATP
C 6H 12O 6 - > 2 C 3H 3O 4O 3 + 2 ATP + 2 NADH + 2 H +
NADH, ou NAD + em seu estado desprotonado (nicotinamida adenina dinucleotídeo), é um so- chamado portador de elétrons de alta energia e intermediário em muitas reações celulares envolvidas na liberação de energia. Observe duas coisas aqui: uma é que a glicólise sozinha não é tão eficiente na liberação de ATP quanto a respiração aeróbica completa, na qual o piruvato produzido na glicólise entra no ciclo de Krebs a caminho dos átomos de carbono que aterram na cadeia de transporte de elétrons. Enquanto a glicólise ocorre no citoplasma, as reações subsequentes da respiração aeróbica ocorrem nas organelas celulares chamadas mitocôndrias.
Glicólise: etapas iniciais
Glicose, que contém uma estrutura de seis anéis que inclui cinco átomos de carbono e um átomo de oxigênio, é transportado para a célula através da membrana plasmática por proteínas especializadas de transporte. Uma vez dentro, é imediatamente fosforilado, isto é, um grupo fosfato é anexado a ele. Isso faz duas coisas: fornece à molécula uma carga negativa, prendendo-a na célula (moléculas carregadas não podem atravessar prontamente a membrana plasmática) e desestabiliza a molécula, configurando para mim mais realidade dividida em componentes menores. < A nova molécula é chamada glicose-6-fosfato (G-6-P), pois o grupo fosfato está ligado ao átomo de carbono número 6 da glicose (o único que fica fora da estrutura do anel). A enzima que catalisa essa reação é uma hexoquinase; "hex-" é o prefixo grego para "seis" (como em "açúcar com seis carbonos") e quinases são enzimas que roubam um grupo fosfato de uma molécula e o fixam em outro lugar; nesse caso, o fosfato é retirado do ATP, deixando o ADP (adenosina difosfato) em seu rastro.
Glicólise: etapas finais
Na sexta etapa, GAP é convertido em 1,3-bisfosfoglicerato (1,3-BPG) sob a influência da enzima pela gliceraldeído 3-fosfato desidrogenase. As enzimas desidrogenase fazem exatamente o que seus nomes sugerem - elas removem átomos de hidrogênio (ou prótons, se você preferir). O hidrogênio liberado do GAP encontra o caminho para uma molécula de NAD +, produzindo NADH. Lembre-se de que, começando com esta etapa, para fins de contabilidade, tudo é multiplicado por dois, uma vez que a molécula inicial de glicose se torna duas moléculas de GAP. Assim, após esta etapa, duas moléculas de NAD + foram reduzidas a duas moléculas de NADH.
A reversão de fato das reações de fosforilação anteriores da glicólise começa no sétimo passo. Aqui, a enzima fosfoglicerato quinase remove um fosfato de 1,3-BPG para produzir 3-fosfoglicerato (3-PG), com o desembarque de fosfato em ADP para formar ATP. Como, novamente, isso envolve duas moléculas de 1,3-BOG para cada molécula de glicose que entra na glicólise a montante, isso significa que dois ATPs são produzidos em geral, cancelando os dois ATPs investidos nas etapas um e três.
Considerações energéticas e o destino do piruvato
De qualquer forma, a maior parte do piruvato produzido na glicólise se move para a matriz mitocondrial (análoga ao citoplasma de células inteiras) e entra no ciclo de Krebs, também chamado ciclo do ácido cítrico ou ciclo do ácido tricarboxílico. Essa série de reações serve principalmente para gerar muitos portadores de elétrons de alta energia, tanto o NADH quanto um composto relacionado chamado FADH 2, mas também produz dois ATP por molécula de glicose original. Essas moléculas migram para a membrana mitocondrial e participam das reações em cadeia do transporte de elétrons que liberam mais 34 ATP. Na ausência de oxigênio suficiente (como quando você está se exercitando vigorosamente), parte do piruvato sofre fermentação, um tipo de metabolismo anaeróbico no qual o piruvato é convertido em ácido lático, gerando mais NAD + para uso em processos metabólicos.