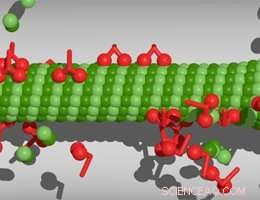
Motores moleculares caminhando ao longo de um microtúbulo. Crédito:M. Rank / PRL
Polímeros filamentosos chamados microtúbulos desempenham papéis vitais na segregação cromossômica e transporte molecular. Uma equipe da LMU agora examinou como os comprimentos dos microtúbulos variam em resposta às mudanças na disponibilidade de seus componentes protéicos.
As células possuem um esqueleto interno, o que permite que eles alterem sua forma e migrem ativamente. Este citoesqueleto é composto por vários sistemas de filamentos, dos quais os microtúbulos são um. Os microtúbulos cilíndricos têm paredes compostas por 13 protofilamentos, cada uma consistindo em subunidades heterodiméricas contendo duas proteínas tubulinas relacionadas. Os microtúbulos não apenas conferem estabilidade mecânica às células e ditam suas formas, eles também servem como uma rede de transporte intracelular. Além disso, microtúbulos são os principais constituintes do fuso mitótico, que medeia a segregação ordenada dos conjuntos de cromossomos replicados nas duas células filhas durante a divisão celular. Todas essas funções requerem regulação dinâmica dos comprimentos dos microtúbulos. Um grupo de físicos LMU liderados pelo Professor Erwin Frey, em colaboração com o professor Stefan Diez (Universidade Técnica de Dresden e Instituto Max Planck de Biologia Celular e Molecular e Genética, Dresden), desenvolveram agora um modelo no qual as proteínas motoras responsáveis pelo transporte de carga ao longo de protofilamentos também servem para regular o comprimento dos microtúbulos. O modelo é descrito e validado experimentalmente na revista. Cartas de revisão física .
Em trabalhos anteriores, O grupo de Frey mostrou que a densidade dos motores moleculares ligados aos filamentos tem um impacto sobre se o microtúbulo cresce ou encolhe, e que seu efeito depende do comprimento do filamento em questão. Quanto mais comprido o microtúbulo, quanto maior o número de proteínas motoras que ele pode acomodar. Moléculas motoras chamadas cinesinas procedem ao longo do protofilamento, passando de um dímero para o próximo. Quando uma proteína cinesina chega ao fim, ele se desprende do filamento levando a tubulina à qual está ligado. Consequentemente, se a densidade do motor no protofilamento for alta, o encolhimento continuará. Por outro lado, um novo dímero de tubulina pode se ligar ao final. No fim, o encolhimento dependente do motor, portanto, compete com o crescimento dos microtúbulos. "Portanto, assumindo que os recursos (ou seja, tubulinas e motores moleculares) estão presentes no acesso, haverá um comprimento de filamento no qual as taxas de crescimento e encolhimento se equilibram, "diz Matthias Rank, primeiro autor do estudo. Contudo, em uma célula real, esses componentes provavelmente não estarão disponíveis em quantidades ilimitadas. Por exemplo, a formação do fuso mitótico esgota significativamente o número de moléculas de tubulina livres na fase solúvel do citoplasma. No novo estudo, os pesquisadores exploraram os efeitos dessa limitação de recursos na regulação do comprimento dos microtúbulos.
Usando simulações baseadas em um modelo matemático de dinâmica de polímeros, eles descobriram que, nessas condições, dois mecanismos distintos de regulação do comprimento entram em ação. Qual destes se torna dominante depende das concentrações relativas das tubulinas e das proteínas motoras:Em uma certa faixa de concentração, o equilíbrio dinâmico entre crescimento e encolhimento dos microtúbulos opera como se os recursos não fossem limitantes. “Mas as coisas são diferentes quando um dos recursos necessários está em falta”, diz Rank. "Esse é o caso, por exemplo, quando não há moléculas motoras suficientes disponíveis para desencadear a despolimerização rápida dos protofilamentos. "Nesta situação, os microtúbulos continuam a crescer até que a concentração de tubulinas caia abaixo de um valor crítico. Além disso, existe uma faixa de concentração em que ambos os processos estão ativos. "Nesse caso, observamos que os microtúbulos vêm em dois tamanhos e que às vezes alternam entre os dois comprimentos ", disse Frey. “Em termos físicos, isso pode ser descrito como uma transição de fase. "Experimentos in vitro realizados por seu coautor em Dresden confirmaram a existência desse regime de transição previsto pelo modelo de Munique. A equipe está convencida de que seus resultados também são aplicáveis a outros sistemas poliméricos , e eles suspeitam que a limitação de recursos-chave pode desempenhar um papel importante na regulação de outros processos celulares.